



Что произошло с эволюцией животного мира после Кембрийского взрыва? Взгляд биотехнолога
 2664
2664 0
0
К настоящему времени накапливается все больше данных о том, что ход эволюции биоты на Земле происходил в различные эпохи с разной скоростью. Особенно выделяют Кембрийский период (примерно с 570 по 500 млн. лет назад), который характеризовался колоссальным увеличением разнообразия всего живого. Современные палеонтологические данные свидетельствуют о том, что ни до, ни после Кембрийского периода не было такого бурного развития эволюционных изменений, такого увеличения численности таксонов, появления многих принципиально новых таксонов. Закономерно, что этот период отмечен определением Кембрийский взрыв. Кембрийский эволюционный взрыв – одна из величайших загадок в истории развития жизни на Земле. Многие исследователи задаются вопросом, с чем связано происхождение Кембрийского взрыва?
Ответ на этот вопрос, на мой взгляд, могли бы получить российские исследователи А.Марков, В. Анисимов, А. Коротаев, опубликовавшие в 2010 г. в Палеонтологическом журнале очень интересное исследование «Взаимосвязь размера генома и сложности организма в эволюционном ряду от прокариот к млекопитающим» [1]. В этой работе были проанализированы изменения величины минимального размера генома (МРГ) в эволюционной линии из нескольких видов животных, находящихся на разных уровнях эволюционного развития. Авторы выбрали из надежных литературных источников данные по геномике девяти видов животных (табл.1).
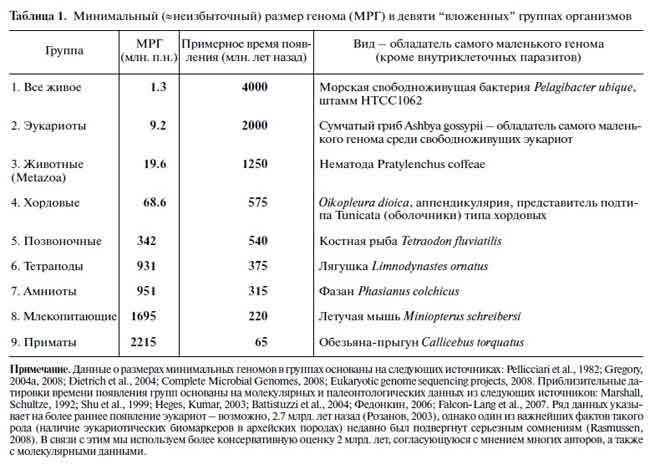
[Марков с соавт., 2010]
Затем ученые совершенно логично провели сравнение нескольких математических моделей, потенциально применимых в данном анализе. Они оценивали два семейства двухпараметрических моделей – экспоненциальные и гиперболические, а также трехпараметрические – степенную экспоненциальную, степенную гиперболическую и двухэкспоненциальную модели. Оптимальные параметры моделей, отражаемых плавными кривыми, подбирались прямым численным перебором при помощи метода наименьших квадратов. В итоге было показано, что в данной эволюционной линии (из девяти организмов, относящихся к различным таксонам) наблюдаемый рост МРГ лучше всего описывается моделью двухэкспоненциальной разновидности гиперэкспоненциального роста (рис.1). Авторы отмечают, что «замедление роста МРГ после появления млекопитающих не означает, что столь же сильно замедлился и рост сложности организмов», однако это утверждение сделано, скорее, на основании общих знаний, чем по результатам оценки параметров их собственной двухэкспоненциальной модели. Соответственно, по результатам анализа данных в статье делается заключение – «эволюция биосферы и социума по пути к общему усложнению является самоускоряющимся (автокаталитическим) процессом» [1]. Я полагаю, что данный вывод совершенно обоснован, но не универсален, то есть он применим только к начальному периоду эволюции на Земле – только к докембрийскому периоду. Более того, в фанерозое характер эволюции животного мира принципиально изменяется. Попробую доказать эти утверждения основываясь на данных А.Маркова с соавторами.
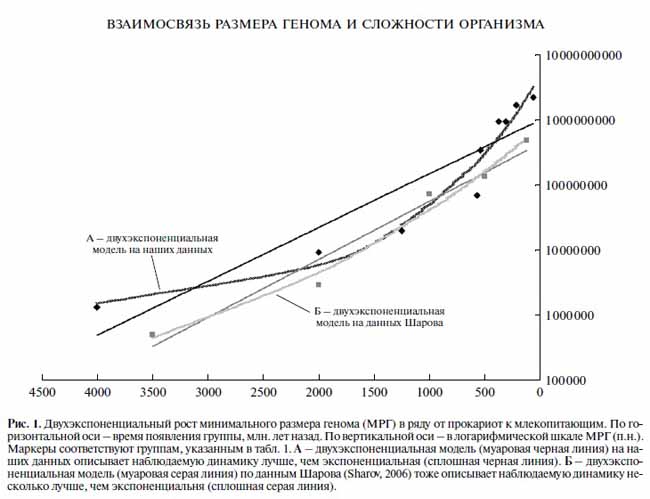
[Марков с соавт., 2010]
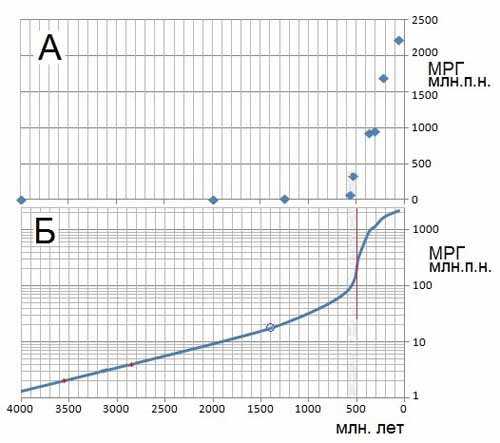
Рис.2. Графический анализ данных, иллюстрирующих увеличение минимального
размера генома (МРГ) девяти видов от прокариотов до млекопитающих.
А – данные, представленные в таблице 1 [Марков с соавт., 2010].
Б – модель характера роста МРГ, отображаемая S-образной кривой,
имеющей точку перегиба, представленная в полулогарифмическом
масштабе. Красной линией отмечен период наивысшей скорости
увеличения МРГ.
Любой биотехнолог с первого взгляда на рисунок 1 узнает в расположении экспериментальных точек S-кривую. Банальную S-кривую, аналогичную тем, что описывают динамику роста численности популяции, например, бактерий в жидкой ростовой среде в замкнутом объеме (рис. 2). Остановимся, тем не менее, на описании такого типа процессов более подробно, поскольку в дальнейшем мы будем постоянно сопоставлять «плавную кривую МРГ» и «биотехнологическую» S-кривую.
Напомню, рост бактерий, внесенных в стерильную ростовую среду, начинается с деления нескольких первых клеток (лаг-фаза роста) и продолжается все более массовым делением присутствующих в колбе. Когда в деление вступают все клетки культуры, рост их числа начинает увеличиваться экспоненциально – в самом быстром темпе (экспоненциальная фаза роста). Если в ростовую среду не вносят необходимых добавок, в какой-то момент клетки начинают ощущать недостаток ростовых субстратов, а затем и испытывать голод. Скорость роста культуры постепенно снижается до тех пор, пока численность клеток не становится постоянной – «выходит на плато». Динамика снижения скорости роста часто аналогична динамике начала роста: несколько клеток ощутили нехватку субстратов и не разделились с обычной скоростью, затем таких клеток становится все больше и больше. Практически так же происходит развитие с последующим торможением роста культуры (отражаемое S-кривой) в случаях накопления в замкнутой среде токсичных продуктов жизнедеятельности.
Итак, точки на кривой МРГ я рассматриваю как точки на S-кривой, то есть на кривой, имеющей некую точку перегиба. На основании данных, приведенных в таблице 1, с помощью программы Excel можно построить графики, как в линейных (рис. 2 А), так и в полулогарифмических координатах (рис. 2 Б). Оба наши графика используем для проведения анализа динамики изменений минимального размера генома в ходе эволюции животного мира. Прежде всего, проведем дополнительные построения на рисунке 2 Б – построим касательную к кривой в точке перегиба, и тем самым разграничим участок (временной интервал) экспоненциального роста МРГ и участок роста МРГ в режиме торможения. Ясно, что момент перегиба, отмеченный нами на графике красной прямой, соответствует экстремуму, в данном случае – временнόму интервалу, характеризующемуся максимальной скоростью увеличения МРГ. После перегиба процесс увеличения размера геномов продолжается, но при этом начинается торможение темпа увеличения МРГ, что, на мой взгляд, требует более детального обсуждения и объяснения. Совершенно очевидно, что выявленная точка перегиба с разумной степенью уверенности соответствует периоду Кембрийского взрыва, что не противоречит общепринятым взглядам.
Таким образом, резюмируя наши рассуждения, можно утверждать, что динамика изменения МРГ может быть отображена S-образной кривой с точкой перегиба (в момент максимального роста МРГ). На мой взгляд, принципиальная ошибка А.Маркова с соавторами состояла в том, что они при решении задачи рассматривали модели, отображаемые исключительно плавными кривыми.
Что же из этого следует? Полагаю, что к дальнейшему обсуждению полученных выводов стоит относиться достаточно серьезно. Вот несколько моих осторожных соображений по данному поводу.
Прежде всего, с каждой неделей в мире появляется все больше данных по геномике новых и новых представителей самых разных таксонов. Несомненно, стоит попробовать «насытить» анализируемую базу МРГ большим количеством «точек» для более корректного построения S-кривой.
Нет оснований полагать, что по мере снижения темпа увеличения размера генома животных эволюция биоты на Земле со временем остановится. Речь идет, скорее, о переключении механизмов эволюции с простого увеличения объема ДНК на усложнение регуляции и повышение эффективности функционирования имеющихся структур геномов.
Самое интересное продолжение размышлений и исследований данной темы, на мой взгляд – все, что касается момента перегиба S-кривой. Напомню, момент перегиба S-кривой у бактерий определяется началом ощущения дискомфорта от ростовых условий – нехваткой субстратов или избытком продуктов жизнедеятельности. А что конкретно послужило сигналом для начала торможения увеличения МРГ у животных?
Особенно интересно включить в сравнение геномные данные о представителях высших таксонов животных. С развитием лабораторных методов работы с ДНК появляются также возможности в качестве «точек» использовать палеогенетические данные.
Косвенным подтверждением S-образного характера развития геномов животных могли бы стать точные и корректно использованные данные по сопоставлению геномов представителей разных таксонов приматов. Действительно ли (как предсказывает S-кривая) чем выше организм в своем эволюционном развитии, тем меньше он отличается от предыдущего, исходного вида?
Известно, что характер эволюции геномов представителей животных и растительных таксонов существенно отличается. Полагаю, что, анализируя эту разницу, можно понять, какая смысловая или функциональная часть генома животных затрагивается торможением. Возможно, окажется правильным уже первое предположение – в отличие от растений, животные обладают развитыми системами генов, обеспечивающих поведенческие, коммуникативные, информационные и когнитивные функции организмов, столь необходимые свободноживущим особям для продолжения рода, то есть для участия в эволюции. Именно в этих генетических системах именно животных повышение качества, эффективности, организованности работы при снижении объемов собственно ДНК может дать эволюционное преимущество.
Не могу не подчеркнуть, что график на рисунке 2 Б позволяет делать некоторые прогнозы. Так можно оценить время удвоения объема геномов для прокариотических организмов, живших на Земле первые 2500 млн. лет (Архей и Палеозой) как 700 млн. лет. Кроме того, наблюдаемый в неявном виде «минорный» перелом S-кривой (на уровне 1400 млн. лет назад) может указывать на некое ускорение развития геномов животных в раннем Мезозое, что предположительно можно связать с началом эволюции одноклеточных эукариотических организмов.
И последнее. Следует отметить, что торможение в процессе увеличения МРГ (или только части генетических систем у части биоты) никоим образом не свидетельствует об отсутствии микроэволюционных процессов в любых таксонах.
В современных условиях нашей планеты, похоже, системы генов поведенческих, коммуникативных, информационных и когнитивных функций у высших животных организмов достаточно хороши, чтобы обеспечивать выживание большинства современных видов. Однако в случае катастроф планетарного масштаба, возможно, понадобятся иные функции и, соответственно, иные генетические системы будут обеспечивать выживание видов. Очевидно, что эти «будущие жизнеобеспечивающие» гены уже есть у представителей каких-либо таксонов. Очевидно также, что для выживания в новых условиях для животных наиболее важно успешное функционирование именно поведенческих, коммуникативных, информационных и когнитивных функций чтобы реализовывать возможности продолжения рода.
Литература:
1. Марков А.В., Анисимов В.А., Коротаев А.В. Взаимосвязь размера генома и сложности организма в эволюционном ряду от прокариот к млекопитающим. Палеонтологический журнал, 2010, №4, 3-14.
Статью подготовил Д.А.Складнев, главный научный сотрудник Института Микробиологии им. Виноградского РАН, доктор биологических наук, профессор
